{kind=link}
Uma das questões básicas em ecologia é como diferentes espécies se juntam formando uma comunidade. Pensando nisso, a partir de pesquisas ecológicas na primeira metade do século XX, foi formulado o princípio da exclusão competitiva. Este princípio diz que duas espécies que ocupam o mesmo nicho ecológico, e portanto competem pelos mesmos recursos, não podem coexistir: uma tende a excluir a outra. O que acontece é que quando duas espécies competem pelo mesmo recurso limitante, os níveis deste recurso tendem a cair até chegar a níveis insuficientes para uma das espécies, que acaba sendo excluída.
Um exemplo clássico do princípio da exclusão competitiva foi um experimento feito por Gause, décadas atrás. Ele cultivou duas espécies de protozoário em laboratório, Paramecium aurelia e Paramecium caudata, separadamente e em conjunto. Quando cultivadas separadamente, ambas sobrevivem. Quando cultivadas juntas, por competirem pelo mesmo recurso limitante, uma exclui a outra.
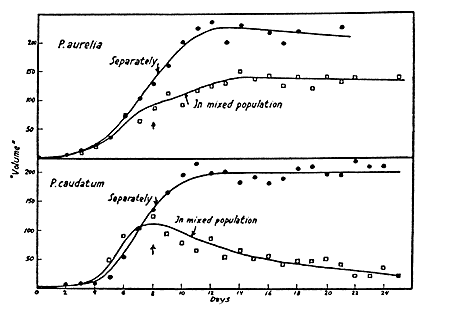
Porém, em 1961, Hutchinson, grande ecólogo, publicou um artigo mostrando uma situação em que o princípio da exclusão competitiva parece não fazer sentido. Ele observou que, no mar e na água doce, há muitas espécies de fitoplâncton. E todas elas competem basicamente pelos mesmos recursos, que são a luz e alguns nutrientes. Como, então há uma diversidade tão grande de espécies competidoras no mesmo ambiente? Por que uma espécie não exclui a outra? A isso, ele chamou de “o paradoxo do plâncton”.

Desde então, surgiram várias hipóteses surgiram para explicar o paradoxo, e até hoje o assunto não está esgotado. O princípio da exclusão competitiva acontece quando o ambiente está em equilíbrio, sem grandes alterações. Isto é fácil de se conseguir em laboratório, mas é difícil de ser encontrado em ambientes naturais. Portanto, a maioria das explicações para o paradoxo do plâncton foca na ausência de equilíbrio.
Ao contrário de experimentos de laboratório, os ambientes naturais são altamente variáveis, tanto no espaço quanto no tempo. A concentração de nutrientes na água, pode amentar ou diminuir em pulsos, e a movimentação da água, por turbulência, ou diferenças de temperatura e densidade, fazem com que os recursos não estejam distribuídos de forma homogênea na água. Apesar destas diferenças de concentração de recursos muitas vezes ocorram em pequena escala, podem ser suficientes para influenciar a dinâmica do microscópico fitoplâncton.
Além disso, em ambientes naturais, há muito mais que nutrientes e fitoplâncton. Organismos zooplanctônicos podem estar constantemente predando organismos das espécies mais abundantes, impedindo que haja um predomínio de uma única espécie. Estes efeitos de predador, além de outros efeitos de teias tróficas, são conhecidos em ecologia por mediar a coexistência de competidores em diferentes comunidades aquáticas e terrestres. Nestes casos, apesar da predação retirar organismos das populações, ela aumenta a diversidade como um todo.
Mesmo sem a predação, a capacidade competitiva não é o único fator envolvido. Muitas espécies de fitoplâncton produzem toxinas. Estas toxinas compensam a desvantagem competitiva das espécies que as produzem, diminuindo o crescimento de espécies competitivamente superiores. E quando há zooplâncton, a produção de toxinas diminui consideravelmente a predação sobre as espécies que as produzem.
Com isso, são múltiplas explicações para o problema levantado por Hutchinson décadas atrás. São tantas explicações, que agora o desafio é saber qual mecanismo dentre tantos, ou qual combinação de mecanismos, é responsável pela coexistência de tantas espécies de fitoplâncton em comunidades reais. Recentemente, este novo desafio foi apelidado de “O paradoxo do paradoxo do plâncton”.
Ciquenta e cinco anos após o clássico artigo de Hutchinson, “The plankton paradox”, ainda há debate sobre o assunto. Isto mostra o quanto a ciência ecológica ainda tem espaço para avançar mesmo em questões básicas.Afinal, a natureza é muito mais complexa, e portanto mais bela, que nossas meras abstrações e modelos matemáticos.
BIBLIOGRAFIA E LEITURAS COMPLEMENTARES
Gause, G.F. (1934) Experimental Analysis of Vito Volterra’S Mathematical Theory of the Struggle for Existence. Science, 79, 16–17.
Hutchinson, G.E. (1961) The Paradox of the Plankton. Am. Nat., 95, 137–145.
Record, N.R., Pershing, A.J., & Maps, F. (2014) The paradox of the “paradox of the plankton.” ICES J. Mar. Sci., 71, 236–240.
Roy, S. & Chattopadhyay, J. (2007) Towards a resolution of “the paradox of the plankton”: A brief overview of the proposed mechanisms. Ecol. Complex., 4, 26–33.
Scheffer, M., Rinaldi, S., Huisman, J., & Weissing, F.J. (2003) Why plankton communities have no equibrium: solutions to the paradox. Hydrobiologia, 491, 9–18.